Embryonic development of birds

Red junglefowl, Gallus gallus [a]
Most studies on embryonic development in birds have been carried out on domestic chickens (Gallus gallus domesticus), and the following description is based on these.
Egg and fertilisaton
Avian eggs generally have a large yolk which consists of concentric spherical layers of white and yellow yolk substance. There is a small germinal disc containing the female pronucleus at the animal pole, and a funnel of white yolk – called the latebra – extends from the germinal disc to the centre of the yolk. A thin vitelline membrane surrounds both yolk and germinal disc.

Figure 1. Section through domestic hen's egg.
Fertilisation of the egg occurs in the oviduct, before the albumen and shell are added to it. The egg is laid about 24 hours after fertilisation, by which time the development has reached the blastula stage.
Cleavage and blastula
Consistent with having a large yolk, cleavage is meroblastic and is restricted to the germinal disc, and cell divisions do not extend into the yolk at all. The first division passes near the centre of the germinal disc and the next few divisions are at right angles to the preceding one, but then divisions becomes more irregular and asymmetric (figure 2).
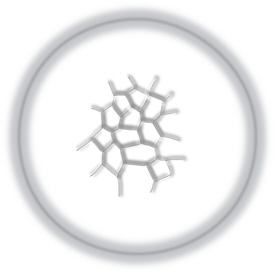
Figure 2. View from above of the germinal disc after the first few cleavages.
Hence, the early divisions gives rise to a disc of cells (blastoderm), several layers thick, sitting on top of the yolk, with a subgerminal cavity in-between. Cells on the periphery of the disc are open to the cytoplasm of the germinal disc, i.e. they form a yolk syncytial layer (YSL).
Across most of the disc, the cells thin to a substantially single layer (but see below) of epithelial cells called the epiblast. This thinning appears to be mainly through cells of the deeper layers being shed, and possibly through some of the lower layer cells being intercalated with the uppermost layer. The central area is relatively translucent and is called the area pellucida, and beneath this is a substantially cell-free subgerminal space. Around the margins of the germinal disc, the cells remain several layers thick and this relatively opaque peripheral ring is called the area opaca.

Figure 3. Longitudinal section through the blastoderm.
The hypoblast (sometimes called the primary hypoblast) then forms. Some of its cells arise as separate ‘islands’ beneath the epiblast; it is not known to what extent these are left-overs from the preceding thinning process or arise by delamination (ingression) from the overlying epiblast. Most of the hypoblast cells arise from the posterior marginal zone (especially the area known as Koller’s sickle, named because it is crescent shaped when viewed from above) by some of its underlying cells spreading anteriorly and incorporating the ‘islands’. The space between the epiblast and hypoblast is the blastocoel.

Figure 4. Longitudinal section through the blastula.
For a fate map of the blastula see here.
Gastrulation
The key feature of gastrulation in birds is the primitive streak.
Initially this is a thickening of the epiblast along its midline, originating close to its posterior end (just forward of Koller’s sickle) and then extending anteriorly until it reaches a maximum extent of about 2/3 across the area pellucida. As this thickening progresses along the epiblast, in tandem with it, a lower layer of cells spreads from the posterior margin. This endoblast (or secondary hypoblast) displaces the (primary) hypoblast anteriorly (figure 5). When the primitive streak reaches its maximum length a groove develops on its dorsal (upper) surface along its length, culminating in a funnel-shaped depression at its anterior end, known as the primitive pit or Hensen’s node.

Figure 5. Early primitive streak, at it's maximum extent, viewed from above, with the epiblast faded at the margin to view the underlying hypoblast and endoblast.
The epiblast spreads (epiboly); and it is through the primitive groove and pit that epiblast cells now ingress between the epiblast and endoblast. This translocation involves a change from an epithelial nature of the epiblast cells to a mesenchymal nature (epithelial-to-mesenchymal transition, EMT) such that the cells can migrate.

Figure 6. Transverse section through the primitive streak.
- Some cells entering the primitive groove move across the intervening space and enter the endoblast to become endoderm (progressively displacing the endoblast to the sides of the embryo). Because the endoderm is an epithelial tissue, these migrating cells must revert to an epithelial nature i.e. undergo a mesenchymal-to-endothelial transition (MET).
- Other cells spread out to form mesoderm between the overlying epiblast (ectoderm) and underlying endoderm.
In general, early cells entering the primitive streak become endoderm and later ones become mesoderm; but at any particular time some cells entering the streak are presumptive endoderm and some presumptive mesoderm. - Epiblast cells that do not enter the primitive streak remain as the ectoderm.
Early cells entering the primitive pit (Hensen’s node) move anteriorly, enter the endoblast and become the endoderm of the presumptive foregut. Cells following these become head mesenchyme, prechordal plate mesoderm, and then chordamesoderm which becomes the most anterior part of the notochord, i.e. starting at the most anterior, progressively more posterior parts of the mesoderm are laid down. When the posterior-ward growth of the chordamesoderm reaches the primitive pit, the pit itself starts to retreat posteriorly; and, as it moves progressively further posteriorly, cells entering the node become progressively more posterior parts of the chordamesoderm (and subsequently notochord).
While gastrulation is proceeding, the cells of the marginal zone start to spread outwards, beginning the extraembryonic membranes, see below.
Neural tube
The cells of the dorsal ectoderm (anterior to the retreating primitive streak) converge towards the midline and extend along the anterior-posterior axis (convergent extension). Those cells near the midline also thicken by changing from cuboidal to columnar to form the neural plate. The lateral edges of the neural plate begin to rise, on each side forming neural ridges and then folds, which fuse to form the neural tube and re-form the overlying ectoderm.

Figure 7. Transverse section to show formation of the neural tube and somites.
Closure of the neural folds starts at two points:
- at the level of the future midbrain, and
- at the hindbrain-cervical boundary;
and progresses in both directions (anteriorly and posteriorly) from each of these.
The neural tube has closed in the anterior part of the embryo before the primitive streak has completely regressed posteriorly.
Neural crest cells
Neural crest cells arise at the edges or crests of the neural folds just before the neural tube closes (i.e. are archetypal neural crest cells) and then disperse laterally.
Mesoderm
Notochord
Notochord
As indicated above, chordamesoderm along the midline forms the notochord (forming anteriorly before the most posterior chordamesoderm has entered through the primitive streak).
Somites
Either side of the notochord the mesoderm splits into a double layer:
- an outer ‘somatic mesoderm’ which lies within the ectoderm (and subsequently contributes to connective tissue), and
- an inner ‘splanchnic mesoderm’ which overlies the endoderm (and subsequently forms the circulatory system).
Immediately either side of the notochord, what is known as the paraxial mesoderm segments into somites which form as pairs either side of the notochord. These form in tandem with the neural tube; and, as with the notochord and neural tube, because of the rearward progression of the primitive pit, the somites at the anterior of the embryo develop well before those at the posterior e.g. the anterior somites are visible before gastrulation completes.

Figure 8. Beginnings of the neural ridges and somites, viewed from above.
Coelom
The remaining space between the two layers of mesoderm extends all around the embryo to form the body cavity or coelom, and beyond the embryo itself it forms the chorionic cavity, bounded by extraembryonic membranes (see below).
Endoderm / gut
As part of the formation of the extraembryonic membranes (see below) the ventral edges of the embryo grow laterally. These folds include the endoderm, and part of this fuses to form the primitive gut (figure 9).

Figure 9. Transverse section to show early embryonic folds.
Extraembryonic membranes
The folds from the edges of the embryo extend and fuse both above (dorsal) and below (ventral) the embryo.
The dorsal folds include ectoderm and (an extension of) the somatic mesoderm. (Anteriorly this fold is known as the cephalic amniotic fold, and posteriorly as the caudal amniotic fold.) When these folds meet above the embryo they form:
- an inner amniotic cavity (after which the amniotes are named, the amnion is the membrane that surrounds this cavity), this fluid-filled cavity surrounds the embryo; and
- part of an outer chorionic cavity (its membrane is called the chorion) which is lined with extraembryonic mesoderm and in effect is an extension of the embryonic coelom.
Note that although both of these membranes comprise layers of ectoderm and mesoderm, the order of these layers in relation to the cavity they enclose is reversed.

Figure 10. Longitudinal section through extraembryonic membranes,
before fusion of the folds to form the amniotic and chorionic cavities.
The ventral folds comprise an extension of the ectoderm and somatic mesoderm on the outside, with endoderm and splanchnic mesoderm on the inner surface; and when they meet below the embryo:
- the ectoderm and somatic mesoderm complete formation of the chorionic cavity; and
- the endoderm and splanchnic mesoderm surround the yolk, forming the yolk sac.
A further extension of the endoderm and mesoderm, from the hind-gut region, forms an additional cavity called the allantois which collects waste products from the embryo.
After the extraembryonic folds have fused and the membranes are complete, the chorion completely surrounds the embryo and the extraembryonic compartments including the allantois. But as the embryo develops, the yolk shrinks and the allantois grows until it completely surrounds the chorionic cavity (and lies within the egg shell).
Image credits
Graphics are by David Swift unless stated otherwise.
Background image for the page banner is by DrKontogianniIVF from www.needpix.com/photo/download/674083/embryo-ivf-icsi-infertility-fertility-free-pictures-free-photos-free-images-royalty-free.
a. Image of Red Junglefowl by Francesco Veronesi, https://commons.wikimedia.org/wiki/File:Red_Junglefowl_-_Thailand.jpg licensed under the Creative Commons Attribution-Sare Alike 2.0 Generic licence.
Page created October 2020.