Embryonic development of reptiles

Sea turtle [a]
There are four main groups of extant reptiles: turtles, lizards, crocodilians, and snakes. Compared with other classes of vertebrates, embryological studies of reptiles have been limited, with most known about turtles. Although turtles are somewhat atypical because of their exoskeleton (carapace on their back, plastron underneath), their early embryonic development is reasonably representative of reptiles, so I have based the following description on these.
Egg and fertilisation
All turtles are oviparous. They have large yolky eggs, similar to those of birds, except that there is no chalaza (true for all reptile eggs) or air space (some reptile eggs have an air space). There is also less albumin, the shell is generally thinner, and the calcium carbonate content of the shell is present as aragonite (all reptiles) rather than calcite.

Figure 1. Turtle egg.
Fertilisation occurs at the head of the oviduct, before addition of albumin and the shell. Embryonic development has reached gastrulation by the time the egg is laid.
Cleavage and blastula
Consistent with the large yolk, cleavage is meroblastic, and cell divisions are restricted to the germinal disc. Initial divisions are perpendicular to preceding ones, but soon become less synchronised, and repeated cell divisions lead to a disc of small blastomeres on the surface of the yolk.
The blastomeres segregate to form a surface layer of epithelial cells called the epiblast, and a thin underlying layer of cells called the hypoblast, with a very thin intervening blastocoel. In the central region of the epiblast the cells are columnar and form what is called the embryonic shield. This is surrounded by an annular area pellucida which overlies the blastocoel; and around this is the area opaca which overlies the cells of the marginal zone (also called the germ wall). Cells at the periphery of the marginal zone do not divide completely, but their cytoplasm is continuous with the yolk; i.e they form what is called a yolk syncytial layer (YSL). (It is unclear whether at this stage there's a subgerminal space between the hypoblast and the yolk, but there appears to be by the time the blastopore extends through the hypoblast (see below). If there isn't a subgerminal space, then the YSL may extend beneath the hypoblast.)
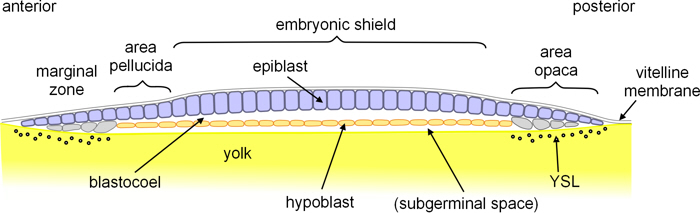
Figure 2. Longitudinal section through turtle blastula.
Gastrulation
Gastrulation begins with a thickening of the posterior edge of the embryonic shield, a thickening which extends across the posterior area pellucida, and is called the blastoporal plate. A transverse invagination then forms near the anterior side of the blastoporal plate, which is the beginning of the blastopore.
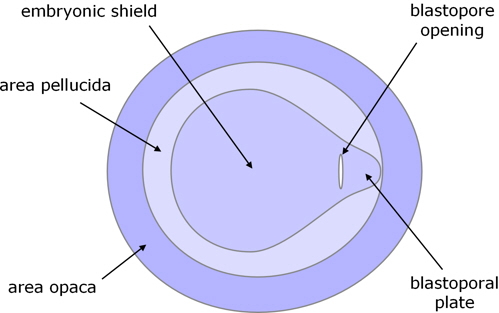
Figure 3. Dorsal view of early blastopore.
Epiblast anterior to the blastopore spreads (epiboly) and cells from this area begin to move (involute) over the anterior lip of the blastopore to form a bilayer structure (figure 4) over the roof of the blastoporal canal which extends anteriorly as well as deeper. Posterior to the blastopore, the epiblast also expands, cells move (involute) into blastoporal canal and contribute to its floor. It appears that some cells in this posterior area also proliferate and enter deeper layers of the blastoporal plate by ingression.
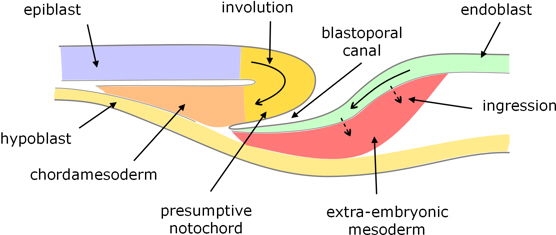
Figure 4. Longitudinal section through the blastoporal canal.
Considerable areas of the epiblast and blastoporal plate converge toward and involute through the blastoporal canal:

Figure 5. Fate map of turtle blastula, viewed from above.
From the epiblast:
- cells involuting along the centreline give rise to chordamesoderm along the roof of the blastoporal canal, some of which develops into the notochord; and
- cells involuting either side of this become paraxial and lateral mesoderm.
From the blastoporal plate:
- cell spreading into the blastopore form the floor of the blastoporal canal, and become endoderm; and
- ingressing cells become predominantly extraembryonic mesoderm.
Cells reaching the ventral surface of the blastula, displace the hypoblast and become endoderm.
The blastoporal canal lengthens anteriorly, and after extending about half-way under the overlying embryonic plate it breaks through into the subgerminal cavity, to form a continuous tube between the dorsal and ventral surfaces. This opening then recedes posteriorly, possibly due to the floor of the blastoporal canal dispersing laterally, and the remaining blastoporal canal shortens such that its ventral opening is only a little anterior to the dorsal opening.
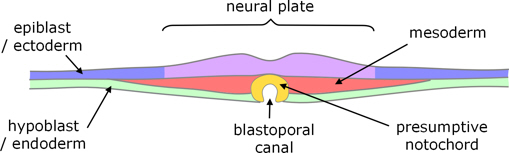
Figure 6. Transverse section, where the blastoporal canal breaks through to the subgerminal cavity.
Neural tube
After the blastoporal canal has formed, the overlying epiblast converges on the midline and thickens to form the neural plate. The lateral edges of the neural plate begin to rise, forming neural ridges and then folds on each side, which fuse to form the neural tube and re-form the overlying ectoderm (figure 7). Fusion starts at the posterior end of the presumptive head and propagates anteriorly and posteriorly.
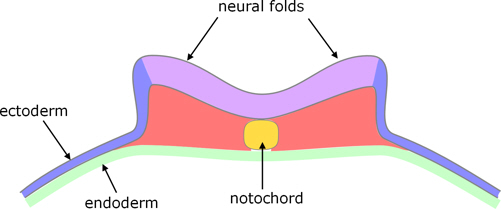
Figure 7. Transverse section, showing early neural folds
(based on a crocodillian rather than a turtle).
Neural rest cells
Neural crest cells (NCCs) form at the crests of the fusing neural folds (i.e. are typical neural crest cells), and disperse in the usual way, i.e. those from the trunk region form e.g. melanocytes, ganglia and neurons (but not bones).
However, in turtles the production of neural crest cells continues for longer than usual, and gives rise to a population of NCCs which are distinct from the early ones, in terms of both character and fate. This second population of NCCs arises from the dorsal side of neural tube, but then migrates to the ventral side of the embryo where they have a key role in the formation of the bones of the plastron (underbody shell). Whereas NCCs from the trunk region do not normally form bones, this second population, although from the trunk region, have characteristics which are similar to cranial NCCs (which form some bones of the head). [2]
Mesoderm
Notochord
As indicated above, chordamesoderm along the midline forms the notochord.
Somites
Either side of the notochord the mesoderm splits into a double layer:
- an outer ‘somatic mesoderm’ which lies within the ectoderm (and subsequently contributes to connective tissue), and
- an inner ‘splanchnic mesoderm’ which overlies the endoderm (and subsequently forms the circulatory system).
Immediately either side of the notochord, what is known as the paraxial mesoderm segments into somites which form as pairs either side of the notochord. These form in tandem with the neural tube.
Coelom
The remaining space between the two layers of mesoderm extends all around the embryo to form the body cavity or coelom and contributes to the extraembryonic membranes (see below).
Endoderm / gut
As part of the formation of the extraembryonic membranes (see below) the ventral edges of the embryo grow laterally. These folds include the endoderm, and part of this fuses to form the primitive gut (figure 8).

Figure 8. Transverse section through turtle embryo to show early embryonic folds.
Extraembryonic membranes
The folds from the edges of the embryo extend and fuse both above (dorsal) and below (ventral) the embryo.
The dorsal folds include the ectoderm and (an extension of) the somatic mesoderm. (Anteriorly this fold is known as the cephalic amniotic fold, and posteriorly as the caudal amniotic fold.) When these folds meet above the embryo they form:
- an inner amniotic cavity (after which the amniotes are named, the amnion is the membrane that surrounds this cavity), this fluid-filled cavity surrounds the embryo; and
- an outer chorionic cavity (its membrane is called the chorion) which is lined with extraembryonic mesoderm and in effect is an extension of the embryonic coelom.

Figure 9. Longitudinal section through extraembryonic membranes,
before fusion of the folds to form the amniotic and chorionic cavities.
Note that this image is based on the formation of extraembryonic membranes in birds, which is thought to be similar to that in reptiles;
I have not found a specifically reptile-based image.
When the ventral folds meet below the embryo:
- the endoderm surrounds the yolk, forming the yolk sac; and
- the mesoderm and ectoderm complete formation of the chorionic cavity.
A further extension of the endoderm and mesoderm, from the hind-gut region, forms an additional cavity called the allantois which collects waste products from the embryo. As the embryo develops, the yolk shrinks and the allantois grows until it completely surrounds the chorionic cavity (and lies within the egg shell).
After the extraembryonic folds have fused and the membranes are complete, the chorion completely surrounds the embryo and the extraembryonic compartments including the allantois.
Notes
1. Frederica Bertocchini, Cantas Alev, Yukiko Nakaya, Guojun Sheng,(2012), A little winning streak: The reptilian-eye view of gastrulation in birds, Development Growth and Regeneration November 2012; doi: 10.1111/dgd.12014
2. Judith Cebra-Thomas, Anne Terrell, Kayla Branyan, Sonal Shah, Ritva Rice, Lin Gyi, Melinda Yin, Yusha Hu, Gulnar Mangat, Jacqueline Simonet, Erin Betters, Scott Gilbert (2013); Late-emerging trunk neural crest cells in turtle embryos generate an osteogenic ectomesenchyme in the plastron, Developmental Dynamics 242:1223-1235
Image credits
Graphics are by David Swift unless stated otherwise.
Background image for the page banner is by DrKontogianniIVF from www.needpix.com/photo/download/674083/embryo-ivf-icsi-infertility-fertility-free-pictures-free-photos-free-images-royalty-free.
a. Sea turtle https://www.rawpixel.com/image/422049/sea-turtle public domain CC0
A turtle glides through the water at the Texas State Aquarium in Corpus Christi. Original image from Carol M. Highsmith’s America, Library of Congress collection. Digitally enhanced by rawpixel.
Page created October 2020.