Embryonic development of amphibians

Xenopus laevis [a]
There are three groups of present-day amphibians:
- urodeles (having a tail) e.g. salamanders;
- anurans (without a tail) e.g. frogs and toads; and
- caecilians (without limbs) so resemble worms or snakes.
Most embryological studies have been on frogs because their development can be observed easily, especially the African clawed frog (Xenopus laevis) because it is easy to stimulate egg laying, and the following description is based mainly on this species.
Egg and fertilisation
Amphibian eggs are generally large, with a substantial quantity of yolk. In Xenopus the cytoplasm is in two distinct parts, inner and outer (or cortical). The inner cytoplasm of the animal hemisphere is darker than that of the vegetal hemisphere; and, corresponding with this, the animal cortical cytoplasm is pigmented, whereas the vegetal cortical cytoplasm is not. In the haploid egg meiosis is halted at metaphase II, and on fertilisation meiosis completes to yield the diploid zygote.
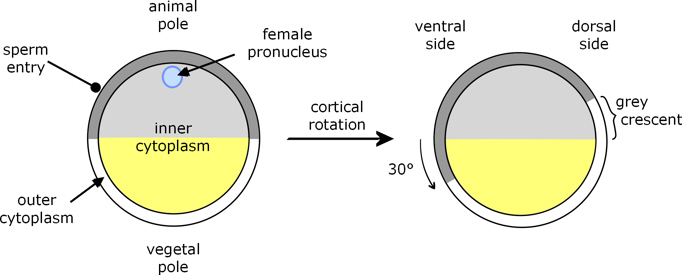
Figure 1. Xenopus fertilisation[a]
Sperm entry and completion of meiosis prompts the cortical cytoplasm to rotate by about 30° relative to the deep cytoplasm, such that part of the underlying animal hemisphere becomes visible as a ‘grey crescent’. Due to this rotation, sperm entry (which always occurs in the animal hemisphere) specifies the dorsal-ventral (back-front) axis of the embryo, because gastrulation begins opposite sperm entry, on the same side as the grey crescent.
Cleavage and Blastula
Cleavage is holoblastic: the first divisions extend right through the zygote. The first cell division goes through the poles of the zygote (meridional), as does the second but at right angles to the first, to produce four approximately symmetrical cells. The third division is at right angles to the first two (latitudinal), and approximately equatorial except that it is somewhat towards the animal pole. This occurs because, although cleavage is holoblastic, division is slower through the yolky vegetal hemisphere; and this pattern continues with subsequent divisions being more frequent in the animal hemisphere, which leads to cells there being smaller than in the vegetal hemisphere.

Figure 2. Xenopus cleavage: the first 3 cell divisions.
A blastocoel begins to develop from the first cell division, and is evident from the 8-cell stage. Eventually it occupies a large part of the animal half of the blastula. Its dome-like roof is formed by numerous small, pigmented micromeres, whilst the vegetal half is composed of large yolk-laden, pale macromeres. Also, the cells of the animal hemisphere and upper part of the vegetal hemisphere form an outer epithelial layer and an inner or deep layer whose cells are more mesenchymal in nature.

Figure 3. Section through Xenopus blastula.
Gastrulation
Before describing the cell movements that occur during gastrulation it’s probably helpful to summarise the fates of different parts of the blastula.

Figure 4. Section through Xenopus blastula section, showing prospective fates following gastrulation.
The legend applies to the remaining figures on this page as well.
- The cells of the animal hemisphere spread (epiboly) to cover the whole of the embryo, to form the ectoderm. Cells along the dorsal centreline form the neural plate, which produces the neural tube and then reforms the epidermis above the neural tube.
- A band of cells (the involuting marginal zone, IMZ) around the equator of the blastula moves inwards and then upwards. Mainly, these form mesoderm, the most dorsal part of which is the notochord.
- Most of the cells of the vegetal hemisphere are enveloped by the expanding ectoderm, are incorporated within the body of the embryo, and develop into endoderm.
The first external sign of gastrulation is when cells just below the equator, called bottle cells, on the dorsal side invaginate to form a crescent-shaped groove, which is the beginning of the blastopore.
However, before this, some deep cells (presumptive prechordal plate) of the dorsal side of the vegetal hemisphere begin to move toward the animal pole. They move close to the inside of the animal dome, but separated from it by a small gap known as the Cleft of Brachet.
Bottle cells. Bottle cells occur on the outer surface of the embryo where invagination occurs. Their outer surface constricts, and this constriction creates a local depression which develops into the invagination through which cells involute. They are called 'bottle cells' because the constriction makes them somewhat bottle-shaped.
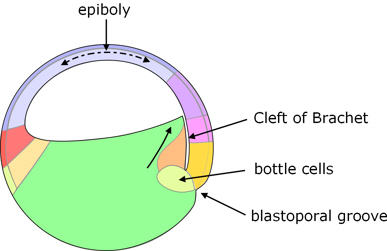
Figure 5. Start of gastrulation.
At the same time, the animal hemisphere begins epiboly (partly by flattening of its cells, and partly by intercalation of cells from different layers) and, as it does so, its cells along the upper edge (dorsal lip) of the blastopore start to roll over the lip (involute) into the blastopore. These cells, which are predominantly presumptive notochord, move towards the animal pole, close to the overlying outer layer of cells, behind the presumptive prechordal plate.
The dorsal part of the blastopore deepens to form a pouch known as the archenteron which gets progressively larger, gradually displacing the blastocoel which in due course almost disappears.
The blastoporal groove progressively extends laterally on both sides until the two ends of the groove meet on the ventral side of the blastula, and the resulting blastoporal groove completely encircles the vegetal pole of the blastula. At the ventral side of the blastopore, cells of the expanding animal hemisphere involute and move towards the animal pole. What were part of the outer layer of cells becomes endoderm, and the deep cells become predominantly mesoderm.
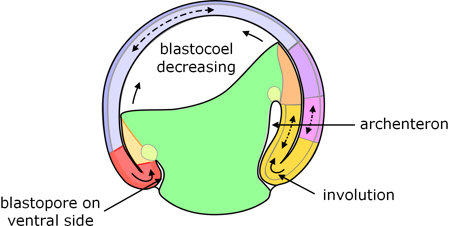
Figure 6. Mid-gastrulation: the archenteron has formed, the blastopore encircles the vegetal pole, and involution is taking place all around the lip of the blastopore.
It will be apparent from the foregoing that involution begins at the dorsal side before the ventral side. Consistent with this, the involuted cells on the dorsal side move further than those on the ventral side, and there is an overall rotation of the inner cells with respect to the outer cell layers.
Endoderm and gut
As epiboly of the animal hemisphere continues, the blastopore becomes smaller, and the cells of the vegetal hemisphere are progressively incorporated into the gastrula where they develop into endoderm. There is a consequent constriction of the blastopore, and reduction of the exposed cells of the vegetal hemisphere until only a small blastopore remains, called the yolk plug. Eventually, the archenteron becomes the digestive tract, with the blastopore as the anus.
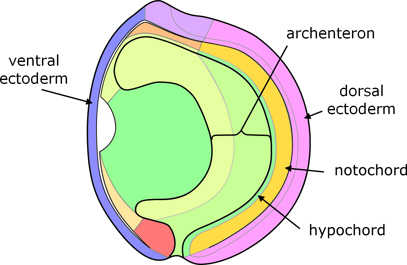
Figure 7. End of gastrulation.
Mesoderm
Notochord
As indicated in figure 4, most of the involuting cells (IMZ) become mesoderm, extending around the embryo between the ectoderm and endoderm. Those along the dorsal midline become notochord.
Coelum
The remaining ‘paraxial’ mesoderm on each side of the notochord forms into a double layer:
- an outer ‘somatic mesoderm’ which lies within the ectoderm (and subsequently contributes to connective tissue), and
- an inner ‘splanchnic mesoderm’ which overlies the endoderm (and subsequently forms the circulatory system).
The space between them extends all around the embryo to form the body cavity or coelum.

Figure 8. Transverse section through the dorsal side after gastrulation.
Somites
Close to the notochord the two layers of mesoderm are called dermatome and myotome, which are joined immediately alongside the notochord (the split that forms the coelum does not extend right up to the notochord, figure 8). The (inner) myotome segments into somites, which arise in pairs either side of the notochord, developing in a regular sequence, in the anterior to posterior direction. The somites arise from the myotome (figures 8 to 12); and in the course of segmentation, the cells of the myotome rotate through 90° which separates them from the dermatome which remains as a sheet between the newly-formed somites and overlying ectoderm (figure 12).
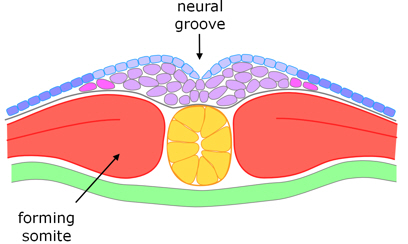
Figure 9. Transverse section through the dorsal side at the beginning of somitogenesis and neurulation.
Neural tube
The neural tube forms during the time when the somites are developing (figures 8 to 12).
On completion of gastrulation, the dorsal surface (ectoderm) comprises a double layer of cells – deep and superficial. This includes the neural plate, although at first there is no clear morphological boundary between the neural plate (which will be internalised as the neural tube) and the presumptive epidermis (which remains on the exterior).
The neural plate narrows and lengthens (through what is called convergent extension) such that both layers of cells converge towards the midline – cells of the upper layer intercalating with the lower cells – to produce a bulge of cells between the notochord and neural groove (figures 9 and 10).

Figure 10. Transverse section through the dorsal side shortly before the sides of the neural groove fuse.
The sides of the groove fuse, almost simultaneously along the full length of the neural groove, usually enclosing a small lumen, although its surrounding cells are mesenchymal rather than epithelial.
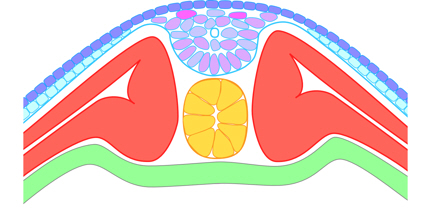
Figure 11. Transverse section through the dorsal side after the neural groove has fused to form the neural keel,
and the somites have almost developed from myotome.
Immediately after this fusion, the cells between the notochord and overlying ectoderm appear disorganised. However, they gradually rearrange /organise into a clearly defined neural tube, involving intercalation and a change from mesenchymal to epithelial cells, this process starting at the ventral (lower) side of the tube, and progressing to the dorsal (upper) side.
As the neural tube forms, a deep layer of dorsal epidermis reforms over the neural tube (and below the superficial epidermis).

Figure 12. Transverse section through the dorsal side after the neural tube and somites have formed.
Neural crest cells
The presumptive neural crest cells arise in the lower layer of cells near the lateral boundary of the neural plate (figure 8). At the time of fusion of the two sides of the neural groove, the forming neural crest cells are not near the point of fusion but in the deep layer, somewhat distant from the midline (figure 10). That is, although there are neural folds, neural crest cells do not originate from the fusion points (which might be thought of as crests) of these folds, either in terms of depth or distance from the midline. These cells migrate (through the tissue, not by gross movement of cells) toward the midline as the neural tube forms, and become associated with the dorsal side of the neural tube, before migrating elsewhere.
Notes
1. R. Keller, D. Shook (2004), Gastrulation in amphibians, Chapter 13 in C. Stern (Ed.) Gastrulation: From cells to embryos, Cold Spring Harbor Laboratory Press.
2. Louie Hamilton (1969); The formation of somites in Xenopus, J. Embryol. exp. Morph 22(2) pp253-64.
3. L. Davidson, R. Keller (1999), Neural tube closure in Xenopus laevis involves medial migration, directed protrusive activity, cell intercalation and convergent extension, Development 126 pp4547-4556.
Image credits
Graphics are by David Swift unless stated otherwise.
Figures 4 to 7 are based mainly on Keller and Shook [1].
Figures 8 to 12 are based mainly on Hamilton [2] for somitogenesis, and Davidson and Keller [3] for neurulation.
Background image for the page banner is by DrKontogianniIVF from www.needpix.com/photo/download/674083/embryo-ivf-icsi-infertility-fertility-free-pictures-free-photos-free-images-royalty-free.
a. Xenopus laevis from https://www.flickr.com/photos/47745688@N05/14808441560 license https://creativecommons.org/licenses/by-nd/2.0/
Page created October 2020.