DNA Polymerase III
DNA Polymerase III holoenzyme
As outlined in replication of DNA, the principal (replicative) enzyme that synthesises DNA in prokaryotes (bacteria) is part of a large complex which is known as the DNA polymerase III holoenzyme, which contains 10 different proteins (12 if the helicase and primases are included), some in multiple copies.
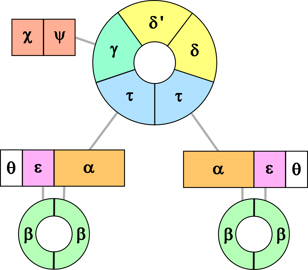
Figure 1. Schematic of the
Pol III holoenzyme.
The holoenzyme comprises three main parts (Fig. 1):
- the polymerase (2 or 3 copies*), each comprising:
- α subunit, the largest component, which adds nucleotides to DNA,
- ε which carries out error correction, and
- θ which assists error correction;
- sliding clamp (2 identical β subunits) – associated with each active polymerase; and
- clamp loader (6 different subunits – δ, δ', γ, τ, χ and ψ) – there are two copies of the τ protein which link the polymerases to the clamp loader (and the helicase, not shown).
*See my note here on whether there are 2 or 3 copies of Pol III. If there are 3 copies then the γ subunit is replaced by a third τ subunit.
Here I will focus on the principal component, the α subunit.
The alpha subunit of DNA polymerase III (Pol IIIα)
Structure of Pol IIIα
The Pol IIIα protein is 1160 amino acids long.

Figure 2. Domains of Pol IIIα. [a]

Figure 2. Domains of Pol IIIα. [a]
It folds into several distinct domains, with substantially distinct functions (Fig. 2). [1]
- Like all DNA polymerases, the section that carries out the principal catalytic function of adding nucleotides to DNA folds into a shape that resembles a cupped right hand, with ‘palm’, ‘thumb’ and four ‘finger’ domains (see Fig. 3).
- To one side of this (at the N-terminal end of the amino acid sequence) is a ‘PHP’ domain which may have a role in proofreading or error-correction.
- To the other side of the ‘hand’ is a domain that binds the sliding clamp (β subunits) which keeps the polymerase in contact with the DNA and greatly enhances the speed and continuity of nucleotide addition.
- Then comes an ‘OB fold’ which typically binds single strand DNA. However, there is some doubt about its function as it seems too far from the single strand DNA entering the polymerase, although it may have a role in sensing the end of an Okazaki fragment and triggering release of the polymerase. [2]
- At the C-terminal end of the amino acid sequence is the domain that binds the τ subunit which links the Pol IIIα to the clamp loader (and to the helicase, DnaB).
How Pol IIIα engages DNA
When Pol IIIα engages DNA, the first 9 base pairs of double strand DNA (from the end of the primer or complementary strand) fit into a cleft formed by the palm, thumb and fingers domains. The thumb and fingers move to wrap partially around the DNA, and hold it in the correct position with respect to the various amino acid side chains of the active site (see below).
Contacts with the DNA include: [3]
- The thumb domain includes a loop (464–470 in the amino acid sequence) which is inserted into the major groove of the DNA towards the active site.
- Several positive side chains (K439, R443, R447, R506) within the thumb domain which provide electrostatic interactions with the negatively charged sugar-phosphate backbone of the DNA.
- As the double strand DNA emerges from the cleft, it is guided, especially by the ‘little finger’ which has several positive amino acid side chains (K871, K872, R876, R877, K881) which attract the negatively charged backbone of the DNA, into the sliding clamp.
- The orifice of the sliding clamp is lined with various amino acids which have non-specific contacts with the DNA and provide an electrostatic cushion for the DNA to pass through.
Overall, approximately 17 base pairs of DNA are in contact with the Pol IIIα, and a further 5 base pairs passing through the sliding clamp.

Figure 3. A 3-D representation of part of DNA Polymerase III. It includes all of the alpha subunit except for the tau-binding domain (as this would obstruct the view of the rest of the protein), one half of the sliding clamp (pale green, the other beta unit is omitted so that the amino acids of alpha that bind it (dark green), and the DNA as it passes through the clamp, can be seen), and the epsilon subunit (purple, note that this appears in two parts; in reality they are linked, but the linking sequence is not included in the crystal structure used for this model). Colouring of the alpha subunit domains follows that used in Fig. 2; the principal amino acids of the active site are red, the loop of the thumb that engages DNA is yellow, and the positive side-chains of the thumb and fingers are pale blue. The DNA is coloured dark purple (template strand) and dark blue (complementary strand). [b]
Contacts with other subunits
Contacts with other subunits include:
- One half of the sliding clamp is bound directly by the Pol IIIα (by amino acids 920-925) into a specific pocket of the clamp.
- The other half is held indirectly via the exonuclease subunit ε which binds Pol IIIα on one side
and the sliding clamp on the other (into an equivalent binding pocket as the Pol IIIα does on the other half).
Note that, as well as its proofreading role, the epsilon subunit greatly enhances the function of the polymerase because of helping to bind it to the sliding clamp. [4] - In addition, the OB domain makes two contacts with the clamp:
by a short helix (1035-1043) which contacts 24-28 on same beta unit as the alpha contact, and
by a loop (1003-13) which contacts 275-278 on the other beta unit (same as the epsilon contacts).
Mechanism for adding nucleotides
Pol IIIα synthesises double strand DNA using deoxynucleoside triphosphates (dNTPs) to build up a complementary strand of DNA base-paired with the template strand. This involves forming a bond between the 3’-OH of the terminal nucleotide on the complementary strand and the α-phosphorus of the incoming dNTP, concomitantly breaking the bond between that phosphorus and the oxygen leading to the β-phosphorus, and hence release of the remaining two phosphates of the dNTP as pyrophosphate (PPi) (see Figs. 4 and 5).

Figure 4. Using a matching dNTP to add a nucleotide to the DNA complementary strand.
However, this reaction by itself is not energetically favourable and would not proceed (there is not enough energy in breaking the bond in the dNTP to form the bond with the 3’-OH, and to compensate for the reduction in entropy by tying the new base into DNA). It has been known for a long time that, in order to make the overall reaction energetically favourable, it is also necessary to split the PPi into two separate phosphates, but not known how this was achieved, especially so as to utilise the energy released. Recent work [5] indicates that splitting of the PPi is an intrinsic part of the polymerase reaction, and suggests that the energy may also be used to move the double strand DNA by one base-pair within Pol IIIα to line up the active site for addition of the next nucleotide.
Active site
These reactions are carried out at Pol IIIα’s active site which is located in the palm domain, between the index finger and thumb. The core of the active site is three aspartate side-chains (at positions 401, 403 and 555 in the amino acid sequence) which are located (in the folded protein) close to the end of the complementary strand and incoming dNTP. [6]
All three aspartates are involved in coordinating two magnesium ions (Mg2+) which are essential for the reaction to proceed; and one of the aspartates (555) also acts as a general base (here meaning the opposite of an acid, rather than a nitrogenous base in DNA) to remove the hydrogen from the 3’-OH of the terminal nucleotide.
One of the Mg2+ ions activates the 3’-OH (lowers its pKa) which means that it more readily loses its hydrogen (to the aspartate), and the electrons freed from the O-H bond are then available to form a bond between the oxygen and the α phosphorus.
This leads to a transition state where the phosphorus is simultaneously bound to five oxygen atoms. The positive charges of both Mg2+ ions help to bind the dNTP to the active site by coordinating the negative charge of its oxygen atoms, and to stabilise this transition state.
Overall, the bond between the oxygen and hydrogen of the 3’-hydroxyl on the terminal nucleotide of the new strand of DNA is transferred to between the oxygen and the α-phosphorus of the incoming dNTP; and, consequently, the electrons of the bond between the α-P and the oxygen between it and the next phosphorus (β) are transferred to the oxygen.
For this reaction to proceed, the dNTP must be aligned closely with the template, the end of the complementary strand, the aspartate at position 555, and the two Mg2+ ions.
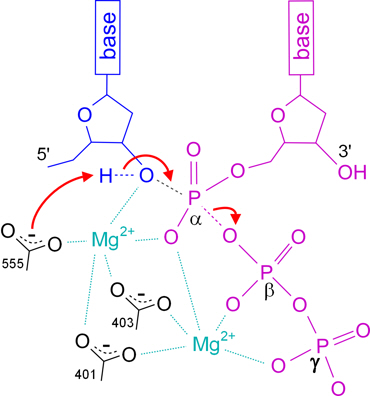
Figure 5. Schematic representation of the Pol IIIα mechanism at its active site.
The terminal nucleotide of the complementary strand is coloured blue, the incoming dNTP is purple, the carboxylate groups of the aspartates are black;
movements of electrons are shown as red arrows, and bonds that are broken or formed are dashed.
The curved dashed lines on the aspartates indicate that the negative charge is spread between the two oxygen atoms.
In addition to the core aspartates, there are several other amino acids that are known to be essential for the polymerase reaction to work.
In the ‘palm’ domain:
- A lysine (positively charged, in position 553, close to one of the catalytic aspartates) forms an electrostatic bridge with the (negatively charged) phosphate of the terminal nucleotide of the complementary strand.
- Two (positively charged) arginines (390 and 396) which coordinate the (negatively charged) phosphates of the incoming dNTP (along with the Mg2+ ions).
In the index finger domain:
- A further two arginines (709 and 710) which, along with the those at 390 and 396, play an important role in targeting the incoming nucleotide to the polymerase active site, and contribute to the catalysis by neutralizing the charge on the triphosphate.
- A tandem glycine-serine (363 and 364) lie in a loop which forms part of the incoming dNTP binding pocket.
- Arising from a detailed comparison of the amino acid sequences from many different DNA polymerases, one study identified a further sequence, from 674 to 682, as essential for the enzyme’s function, and suggested it may be part of the enzyme's active site. [7] However, although these amino acids are in the index finger domain, they seem too far from the known active site to be a part of it, so I have not included them as such in Fig. 3. It may be that they are required for opening and closing of the finger domain (see below).
Binding of dNTP and assembly of the active site
It will be apparent from the above that the active site is in two parts: as well as the key amino acids on the palm which (with the Mg2+ ions) effect the catalysis, there are amino acids on the index finger that also bind the dNTP and are equally important for catalysis to proceed. In the absence of a dNTP, the Pol IIIα adopts an ‘open’ configuration with a space – which includes a dNTP preinsertion site – between the palm and index finger. A dNTP can enter this preinsertion site, and when it does so, in the correct configuration, the enzyme closes (part of the index-finger rotates through about 15°), bringing the dNTP into close proximity with the aspartates and Mg2+ ions, enabling catalysis to proceed. Once the catalysis is complete, the finger opens again to release the residual phosphates, and the Pol III proceeds one more position along the template, vacating the preinsertion site, ready for the procedure to repeat for the next base pair. [8]
Correct base-pairing
The finger closes properly only if the preinsertion site is engaged by a correctly base-paired dNTP (an induced fit).
When an incorrect (non-complementary) dNTP occupies the site, it is not in the right position, so even if the finger tries to close the dNTP is not in the correct position with respect to the aspartates and Mg2+ ions for the catalysis to take place.
This is the main mechanism for how correct base-pairing is achieved.
Selection of correct sugar
As well as identifying the correct dNTP in terms of having the right base, the Pol IIIα must also select the deoxynucleoside (with deoxyribose) rather than a ribonucleoside (with a ribose). It should be noted that ribonucleosides are much more abundant in the cell (perhaps 1000-fold), including adenosine triphosphate (ATP) which is the primary high-energy compound in the cell. The difference between ribose and deoxyribose is that ribose has an additional hydroxyl group, at the 2’ position. Selection for deoxynucleosides is effected by having amino acids at the dNTP presinsertion site that have side chains that occupy the position that would be taken by the 2’-OH, and hence prevent ribonucleosides from successfully entering the preinsertion site. In Pol IIIα the amino acids having this role are thought to be histidine-760 and tyrosine-764. [9]
Proofreading and first-level error-correction
The mechanisms just described achieve a high level of fidelity – having only 1 error in 104 to 106 nucleotides which, bearing in mind the speed of this enzyme of adding up to 1000 nucleotides per second, is astonishing! In addition, some mismatched nucleotides are detected by the enzyme – which may be a role of the loop of the thumb domain that engages the minor groove of the DNA. When this happens, the epsilon subunit (which has exonuclease activity) removes the mismatched nucleotide, and the Pol IIIα moves back one space along the DNA, allowing a further attempt to incorporate a correct nucleotide. This error-correction removes 90-99.9% of mismatches, leaving an overall error rate of only 1 in up to 108. [10]
Within the cell there are further mechanisms for correcting many of the remaining errors.
How could an enzyme such as this evolve?
My purpose in presenting the above description of how Pol IIIα works is of course to prompt and inform the question: How realistic is it for an enzyme such as this to originate in an evolutionary way?
Note that I am not asking, How realistic is it for Pol IIIα or even any of the DNA polymerases that we know of to arise in this way. That is, although all known DNA polymerases have a similar overall ‘right hand’ shape, and employ essentially the same chemical mechanism, I am not suggesting that this must be the only way DNA replication could take place, i.e. that evolution must ‘find’ a (more-or-less) unique solution. On the other hand, from what we know about DNA polymerases we can confidently draw several conclusions about some criteria for a DNA polymerase:
- Any enzyme with DNA polymerase function must have a substantial minimum size of probably at least 200-300 amino acids. Although Pol IIIα is over 1000 amino acids long of which the ‘hand’ part is about 500 amino acids, the core function is carried out by the thumb, first two fingers, and part of the palm between these, which comprise around 200-300 amino acids.
- The crux of the enzyme’s activity is that it must have some means to implement a chemical reaction to add a nucleotide to single strand DNA.
- A further essential criterion for implementing the principal function is that it must be energetically favourable – otherwise it will not proceed.
This sort of minimum size seems reasonable if a protein is to meet the following requirements, which necessarily entail having many specific amino acids in specific 3-D positions in relation to each other, DNA and any other substrates, which requires a 3-D structure of reasonable size in which to locate them.
Again, consistent with my previous comments, although all of the DNA polymerases that we know about use two cations (usually Mg2+) held in place by at least two aspartate side chains (and there are many other essential amino acids involved), I am not suggesting that this is the only way it could be done.
But a DNA polymerase must have some way of doing it. And, just as the DNA polymerases we know about have appropriate amino acids in the right places in their primary sequence so that when the polypeptide is folded they are in the right 3-D position to form the required active site – and this needs to be quite precise – so any DNA polymerase, even one employing a different mechanism, must have the right amino acids in the right positions to form an appropriate active site to effect that mechanism.
And, as mentioned above, in the case of using dNTPs to add nucleotides to DNA, although the essential reaction is only the transfer of a bond from the dNTP to the 3’-OH of the terminal deoxyribose, breaking only the bond with the alpha phosphorous (with release of PPi) does not provide enough energy to form the new bond with DNA. It is necessary concomitantly to hydrolyse the PPi in such a way that the energy released in doing so can be used to enable the polymerase reaction. So far as I am aware at the time of writing, we do not yet know the mechanism for how this is implemented, or which aspects of the polymerase structure are required. But inevitably this energetic criterion imposes further requirements on the enzyme’s amino acid sequence.
While (2) and (3) are at the heart of the enzyme’s activity, as well as requiring assembly of a competent active site, there are several other requirements that must be met.
- A DNA polymerase must be able, not only to bind DNA per se, but to identify and locate itself at the end of its complementary strand so that its active site is positioned appropriately to add a further nucleotide to it.
- Similarly, it must have near that location a suitable recognition site for a dNTP, appropriately positioned to transfer its nucleotide to the end of the complementary strand.
- It must have a means for matching base pairs with a good degree of accuracy.
- It must also be able to discriminate against the preponderance of ribonucleosides in favour of the much less abundant dNTPs.
- A polymerase must be able, after adding a nucleotide, to move along the template DNA to reposition itself for addition of the next nucleotide.
It may be that the sliding clamp and clamp loader are the key players in doing this. But if that is so, it does not make an evolutionary origin any easier, because as more components are involved in implementing a function it compounds the odds against it arising.
The open-close mechanism of DNA polymerases is an effective way of achieving fidelity – allowing the catalysis to proceed only if the right dNTP is in place. No doubt there are other ways this selection could be done; but, again, there is no doubt that this requirement will incur additional essential amino acids, appropriately placed in the primary sequence etc.
Notes
Notes display in the main text when the cursor is on the Note number.
1. Charles McHenry, DNA Polymerase III structure, Molecular Life Sciences, 2014; doi: 10.1007/978-1-4614-6436-5_131-1
2. Roxana Georgescu, Isabel Kurth, Nina Yao, Jelena Stewart, Olga Yurieva and Mike O'Donnell; Mechanism of polymerase collision release from sliding clamps on the lagging strand, EMBO Journal 28, 2981-2991, 2009; doi: 10.1038/emboj.2009.233 .
3. Rafael Fernandez-Leiro, Julian Conrad, Sjors Scheres and Meindert Lamers; cryo-EM structures of the E.coli replicative DNA polymerase reveal its dynamic interactions with the DNA sliding clamp, exonuclease and τ, eLife doi: 10.7554/eLife.11134 .
4. Ana Toste Rego, Andrew Holding, Helen Kent and meindert Lamers (2013); Architecture of the Pol III-clamp-exonuclease complex reveals key roles of the exonuclease subunit in processive DNA synthesis and repair, The EMBO Journal 32, 1334-1343.
5. Jithesh Kottur and Deepak Nair; Pyrophosphate hydrolysis is an intrinsic and critical step of the DNA synthesis reaction, Nucleic Acids Research 46(12), 5875-5885 (2018), doi:10.1093/narlgky402 .
6. Richard Wing, Scott Bailey and Thomas Steitz; Insights into the replisome structure of a ternary complex of the DNA polymerase IIIα-subunit, J. Mol. Biol. 382(4): 859-869; doi: 10.1016/j.jmb.2008.07.058.
7. P. Palanivelu; DNA Polymerases: an insight into their active sites and catalytic mechanism, International Journal of Biochemistry Research & Review 3(3): 206-247 (2013).
8. Sylvie Doublié, Michael Sawaya and Tom Ellenberger; An open and closed case for all polymerases, Structure 7:R31-R35 (1999).
9. Scott Bailey, Richard Wing and Thomas Steitz; The structure of T. aquaticus DNA polymerase III is distinct from eukaryotic replicative DNA polymerases, Cell 126: 893-904 (2006), doi: 10.1015/j.cell.2006.07.027
10. Thomas Kunkel; DNA Replication fidelity, Journal of Biological Chemistry 279(17):16895-16898; doi: 10.1074/jbc.R400006200 .
Image credits
Graphics are by David Swift unless otherwise stated.
Background image for the page banner is from https://commons.wikimedia.org/wiki/File:How_proteins_are_made_NSF.jpg and is in the Public Domain.
a. Image by David Swift, based on Figure 1 in [1]; and incorporating data from Meindert Lamers, Roxana Georgescu, Sang-Gyu Lee, Mike O'Donnell and John Kuriyan, Crystal structure of the catalytic α subunit of E. col replicative DNA polymerase III, Cell 126, 881-892, 2006; doi: 10.1016/j.cell2006.07.028.
b. Image created using Jmol: an open-source Java viewer for chemical structures in 3D (http://www.jmol.org/) and crystal structure 5FKV (doi: 10.2210/pdb5fkv/pdb) from the Protein Data Bank.
Page created December 2018.